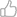两性奥秘 bt两性心理学著作两性霉素说明书雾化两性姿式图片动漫头像
先说结论

先说结论。在现代系统学视野下,真花说(认为单花、两性、花被分离的多心皮类如毛茛和木兰类是被子植物的原始代表)和假花说(原始被子植物的花是具有单性花和具有柔荑花序的)都不完全正确。但假花说错得更离谱,真花说不苛求的话可以认为是正确的。
另外两个学说都已经有一百多岁了啊,已经恍若隔世,现在都没有人提了。两性霉素说明书雾化。。国内的教科书落后得有点夸张。
第二个问题,怎么解释被子植物的系统演化,需要详细说一下。说之前先纠正由于过时的教科书造成的常识性的错误:
3. 由于双子叶植物不是单系群,单子叶植物是单系群,经典的双子叶植物这一概念已经不再适用,但是单子叶植物仍然适用。(单子叶植物出了口恶气:“哈哈本宫为了这么个名份苦苦等了这么多年,终于把双子叶植物这个小婊砸斗死了,善恶终有报两性姿式图片动漫头像,笑看苍天饶过谁两性霉素说明书雾化。”)
先上被子植物大系统研究的最新成果,复旦生科院院长马红实验室去年发在Nature Communications 上面的一篇文章两性心理学著作,用59个核单拷贝基因构建了系统树。先上图:
(Zeng et al., 2014)。菱形代表100%支持率。该研究的开创性意义是,首次得到了金鱼藻科(Ceratophyllaceae,图中紫色部分)和金粟兰科(Chloranthaceae,图中蓝色部分)的高支持率系统位置解决,啃掉了被子植物大系统研究的最后两块硬骨头。这两个科之前的属系关系一直不清楚,这个研究表明他们是姐妹群关系,并成为真双子叶植物的外类群。但是呢,这并不是说这两个科的位置就完全确定了(我没有要钦定,没有任何这个意思!)。分子系统学的问题呢,就在于用不同的分子标记做出来的结果可能会不一样,会有冲突(为什么会这样那就又需要另外开题说了)。这个系统树用到的单拷基因,也是经过”挑选“的,也就是说其实是特意回避了冲突。所以呢,要彻底解决这两个科的位置两性姿式图片动漫头像,还需要更多分子标记,如叶绿体基因组、线粒体基因组系统发育的证据。
但是这个树部分的位置在学界已经达到了比较高的认可度(基部仍然存在变动的可能),可以从下至上依此来看看被子植物的系统发育过程:
最下面黑色的部分是基部被子植物(Basal angiosperms),也被称为ANITA group(用其中五个最具代表性的科的首字母构成)。这是已经被干掉的双子叶植物的一部分尸块。再来看看最最基部是什么植物呢?互叶梅(Amborella trichopoda)。上个图再说:
单目单科单属单种的孓遗植物,只分布在新喀里多尼亚(那地方宝贝多得不得了简直就是地球的藏宝箱你们千万不要告诉别人免得一阵哄抢。)这就是目前发现的被子植物最早分化出去的一支,花的性状也符合真花说两性霉素说明书雾化,基部被子植物里面也以两性、花被分离的多心皮类占大多数,在这个意义上可以认为真花说是正确的。
基部被子植物里面的类群,还有睡莲目(Nymphaeales)。除了大家喜闻乐见的睡莲(另外跟它类似的荷花在离它远得不知道哪里去了的地方大家可以自己找找,我们是强行亲戚多年的陌生人啊)以外,这里面值得注意的还有一个只分布在澳大利亚和印度的小科——排水草科(Hydatellaceae),很丑就不上图了。排水草科本来长期被放在单子叶植物里面,近十年才被人(Saarela et al., 2007,此人很帅很哦)拎出来扔到基部被子植物里面,算是近十年来最大的被子植物系统学变动。(双子叶植物说哈哈老子就算被分尸了也要咬下你单子叶植物一块肉再走)。
然后就是APG II新成立的目——木兰藤目(Austrobaileyales)。中国有的是八角和五味子。
在基部被子植物之上,才分出了单子叶植物。单子叶植物目前是解决良好的单系群。最近在弄单子叶植物,就再深入说说吧:
单子叶植物的最基部是什么植物呢?菖蒲(Acorales)。菖蒲是单子叶植物里面最早分化出去的一支两性心理学著作。
单子叶植物里面还有一个APG充实起来的的新目——露兜树目(Pandales):把以前一些很远的类群搜刮到了一起(我们才是失散多年的亲兄弟啊,露兜树对百部说)。该目各类群形态差异极大,基本归纳不出什么像样的形态共性来,现在姑且就把它们扭在一块儿吧。
跟单子叶植物形成姐妹群的,是双子叶植物的另外一坨尸块。最基部的是木兰类植物(Magnoliids)两性奥秘 bt。可以看见,这个以前被认为是最原始植物的类群,现在离被子植物的基部是如此的遥远。木兰类植物里面有四个目:樟目(Laurales),木兰目(Magnolales),胡椒目(Piperales)是大家熟悉的。在APG下的变动是把木兰目的不产的科林仙科(Winteraceae)和白桂皮科(Canellaceae)扯出来成立了白桂皮目(Canellales),跟胡椒目形成姐妹群两性霉素说明书雾化。
然后就是两个一直位置在真双子叶植物外面游移不定的类群:金鱼藻科和金粟兰科,前面已经提到这里就不详说了。
最上面的,是双子叶植物最大的一团尸块——真双子叶植物(Eudicots),用个”真“跟以前的双子叶植物区分两性奥秘 bt,好像别的尸块都是假的似的两性姿式图片动漫头像。
真双子叶植物呢,也跟被子植物一样,里面搞了一个基部真双子叶植物(你们这样自拥小王国的司马昭之心路人皆知!单子叶植物表示双子叶植物百足之虫死而不僵它还要继续斗下去!)。基部真双子叶植物包括毛茛目(Ranunculales,嘛它在某种意义上还是蛮原始的吧),青风藤科(Sabiaceae),黄杨木(Buxales),昆栏树科(Trochodendraceae),山龙眼目(Proteales两性奥秘 bt两性霉素说明书雾化,嗯前面提到的荷花就在这里。哈哈老娘做不成基部被子植物倒是要弄个基部真双子叶植物当当,睡莲你来打我呀两性姿式图片动漫头像!)
基部真双子叶植物上面呢,又有一个核心真双子叶植物(core eudicots)的概念(喂喂你们是核心我们就是人民群众?你们这样搞小团体会遭报应的基部真双子叶植物很不满诶)。核心真双子叶植物里面呢两性心理学著作,为了说起来顺口,又把最显赫的两个类群编排了两个高大上的名目:蔷薇分支(Rosids)和菊类分支(Asterids),这两个分支里面又分了一些基部类群,然后在之上分别分出有真蔷薇I类分支(常见如豆目,葫芦目,壳斗目,蔷薇目),真蔷薇II类分支(常见如锦葵目,十字花目,无患子目两性奥秘 bt,桃金娘目),真菊I类分支(常见如唇形目,茄目),真菊II类分支(常见如菊目,伞形目),可见真双子叶植物里面真是党派林立两性心理学著作,迟早要完蛋。
被子植物的系统演化差不多就是这样的,很多关系都在现代分子系统学的帮助下被厘清了。现在APGIII中还有大概五个科的位置不清楚,都是一些乏人问津的小科,如茶茱萸科(Icacinaceae),单子叶植物里的多须草科(Dasypogonaceae),但大致应该放在哪一支还是清楚的,最后即便解决了也应该不会对大背景有什么影响。其实还有很多位置确定的类群支持率还不理想,以后可能有变动。这些往往都是寄生植物或者腐生植物(由于这类植物所处环境的选择压力巨大导致基因变异速率太快,分子标记算出来的系统树支持率往往偏低),如霉草科(Triuridaceae,诶我刚得到了很不错的支持率我会到处去乱说么?),还有一些乏人问津的小科,如蒜树科(Huaceae),由于地理偏远难以采到材料,还没有人仔细研究过。
APG系统对传统植物进化理论冲击很大.但有一点,最基部的几个小群过去属于木兰亚纲.所以说真花说还是有一定道理的.但是,传统真花理论认为,最早的花就是花被多,雄蕊多,雌蕊多并且不合生的两性花.但是根据apg系统,最原始的无油樟目,木兰藤目,金粟兰目及睡莲科的排水草科都只有简单的花部结构,并且是单性花常见,反而很多后面的类群还保留了大量传统认为的原始特征比如心皮开放,导管特征原始等等,这就很难自圆其说了.,.事实上,早期的真花说分类系统中,无油樟目,木兰藤目都被放在木兰亚纲较高级的位置.这就能看出分子系统对原有植物形态进化理论的冲击来了.类似的情况不仅出现于被子植物基部,也同样出现在很多后期演化的类群.比如真双子叶植物最基部的毛茛目,其中最先分化的领春木科,单子叶植物最早的菖蒲科在传统分类法中都并不在很原始的位置.
个人理解,apg系统指明植物界的趋同进化远比原先想象的更复杂,很多进步性状是在非常后期的演化中才出现的.最典型的比如导管的进化.早期植物学家就已经知道导管在被子植物几个大类群中可能是各自独立产生并进化的.目前apg系统几乎已经证明了这一点,但比想象中更复杂的是,很多原先认为比较进步的大类群却有形态非常原始的亲戚.比如,菊目被公认是植物界最进化的几个目之一,但是在菊目之后分化出的伞形目,盔瓣花目一个比一个原始,伞形目较原始的几个科和盔瓣花目无论花还是导管都表现出相当原始的特征.这只能说明,菊目(包括和盔瓣花目互为姐妹群的忍冬目的诸多典型合瓣花先进性状都是完全独立产生的.远比传统认知复杂.
顺便介绍个塔赫他间系统对桔梗目起源的说法:桔梗目起源于虎耳草目海桐花亚目.塔赫他间系统的桔梗目实际是现在菊目的基部,而海桐花亚目现在则是伞形目的基部成分.而虎耳草目无论传统分类系统还是apg系统都是作为核心真双子叶植物比较原始的一个基础类群.海桐花亚目早期和虎耳草目混在一起可见其形态上的原始性.
免责声明:本站所有信息均搜集自互联网,并不代表本站观点,本站不对其真实合法性负责。如有信息侵犯了您的权益,请告知,本站将立刻处理。联系QQ:1640731186